|
Арктика и Антарктика
Правильная ссылка на статью:
Руденко О.В., Васильчук А.К., Енина В.В.
Сравнительная характеристика состава субрецентных палиноспектров в донных осадках моря Лаптевых и ледовых комплексах Сибирской Арктики
// Арктика и Антарктика.
2017. № 3.
С. 1-16.
DOI: 10.7256/2453-8922.2017.3.24524 URL: https://nbpublish.com/library_read_article.php?id=24524
Сравнительная характеристика состава субрецентных палиноспектров в донных осадках моря Лаптевых и ледовых комплексах Сибирской Арктики
Руденко Ольга Владимировна
кандидат географических наук
доцент, ФГБОУ ВО "Орловский государственный университет им. И.С. Тургенева"
302026, Россия, Орловская область, г. Орел, ул. Комсомольская, 95
Rudenko Ol'ga Vladimirovna
PhD in Geography
Associate Professor at Orel State University named after I.S. Turgenev
302026, Russia, Orel, ul. Komsomol'skaya, 95

|
olrudenko2011@yandex.ru
|
|
 |
Васильчук Алла Константиновна
доктор географических наук
ведущий научный сотрудник, Московский государственный университет им. М.В. Ломоносова (МГУ)
119991, Россия, г. Москва, ул. Ленинские Горы, ГСП-1, 1, географический факультет, НИЛ геоэкологии Севера
Vasil'chuk Alla Constantinovna
Doctor of Geography
Leading Research Fellow, Laboratory of Geoecology of the Northern Territories, Faculty of Geography, Lomonosov Moscow State University
119991, Russia, g. Moscow, Leninskie Gory, GSP-1, 1,, geograficheskii fakul'tet, NIL geoekologii Severa
|
|
alla-vasilch@yandex.ru
|
|
 |
Другие публикации этого автора
|
|
|
Енина Виктория Викторовна
аспирант, ФГБОУ ВО "Орловский государственный университет им. И.С. Тургенева"
302026, Россия, Орловская область, г. Орел, ул. Комсомольская, 95
Enina Viktoriya Viktorovna
Postgraduate at Orel State University named after I.S. Turgenev
302026, Russia, Orel, ul. Komsomol'skaya, 95
|
|
enina.viktorya@yandex.com
|
|
 |
|
DOI: 10.7256/2453-8922.2017.3.24524
Дата направления статьи в редакцию:
23-10-2017
Дата публикации:
30-10-2017
Аннотация:
Предметом исследования являются вариации субрецентных палиноспектров, по данным изучения ненарушенного поверхностного слоя донных осадков отложений шельфа моря Лаптевых, поднятых мультикорерами в экспедициях ARK 27-3 и Transdrift XXI в 2012-2013 г.г. на шельфе и материковом склоне. В статье обосновывается возможность реконструкции , отражение в маринопалиноспектрах 1) степени ледовитости морского бассейна, индикаторами которой являются наличие и концентрация маркерных видов морских цист динофлагеллат; 2) колебаний речного стока, сигналом которых являются по изменения концентрации в донных осадках органической взвеси терригенного генезиса (споры, пыльца, пресноводные зеленые водоросли), переотложенные в основном из отложений ледового комплекса(едомы); 3) притока трансформированных атлантических вод сигналом для которого является наличие маркерных видов микрофоссилий; 4) климатических условий, определяющих степень продуктивности растений доминирующих биомов побережья, с которого поступает пыльца на шельф моря Лаптевых. Основной метод исследования - маринопалинологический, который включает в себя анализ видового состава и изменений концентрации терригенных и водных палиноморф в донных осадках. Проведен количественный и качественный анализ распределения терригенных и водных палиноморф в донных осадках, отобранных в юго-восточной части моря Лаптевых в затопленной на этапе постгляциальной трансгрессии палеодолине реки Лены. Полученные вариации количественного и качественного состава пыльцы и непыльцевых палиноморф в ненарушенном поверхностном слое донных морских осадков и их сопоставление с данными палинологического анализа из едомного комплекса северо-востока Сибири отложений и субрецентных палиноспектров из северо-восточных районов Сибири позволили оценить современные условия формирования маринопалиноспектров
Ключевые слова:
субрецентные палиноспектры, пыльца, споры, зеленые водоросли, цисты динофлагеллат, донные отложения, едома, море Лаптевых, внутренний шельф, внешний шельф
Исследование выполнено в рамках стипендиальной программы российско-германской Лаборатории по морским и полярным исследованиям им. О.Ю. Шмидта (ААНИИ, г. Санкт-Петербург), и грантов РФФИ №15-05-08497 и РФФИ № 17-05-00794.
Abstract: The research subject is the varieties of subrecent pollen spectra in the unbroken surface layer of bottom sediments of the Laptev Sea shelf, which had been lifted by multicorers in the expeditions ARK 27-3 and Transdrift XXI in 2012-2013 on the shelf and continental slope. The authors substantiate the possibility of reconstruction. The authors consider how the following aspects are reflected in marine pollen spectra: 1) the level of ice coverage of the sea basin, which can be seen in the presence and concentration of index types of marine cysts of dinoflagellates; 2) oscillations of the river flow, assessed by the changes of concentration of organic suspensions of terrigenous genesis in bottom sediments (spores, pollen, fresh-water green algae), transferred mainly from the yedoma sediments; 3) transformed Atlantic water migration, which can traced by index types of microfossils; 4) climate conditions, defining the level of productivity of the flora of the dominating biomes of the shore, pollen to the Laptev Sea shelf comes from. The main research method is the study of marine pollen, which includes the analysis of the species composition and the changes of concentration of terrigenous and water palynomorphs in bottom sediments collected in the southeastern part of the Laptev Sea in the paleovalley of the Lena river, which was drowned during the Postglacial transgression. The obtained varieties of quantitative and qualitative composition of pollen and non-pollen palynomorphs in the unbroken surface layer of bottom sediments and their comparison with the results of the study of pollen from yedoma complex of northeastern Siberia help evaluate the modern conditions of formation of marine pollen spectra.
Keywords: subrecent pollen spectra, pollen, spores, green algae, cysts of dinoflagellates, bottom sediments, yedoma, Laptev Sea, inner shelf, outer shelf
Введение
Палинологические данные являются ценной информацией, которая может быть использована при палеоэкологических реконструкциях и стратиграфическом расчленении позднекайнозойских осадков арктических регионов. Однако применение этого метода для исследования донных осадков арктических морей возможно только с учетом региональных особенностей воздушной и речной транспортировки пыльцы и спор, а также гидродинамических условий морского бассейна. Каждый арктический водоем характеризуется совокупностью условий, в которых происходит захоронение спор и пыльцы в донных осадках: наличием и интенсивностью ледового разноса, направлением господствующих ветров и течения рек, как основных агентов разноса пыльцевого дождя, рельефом морского дна и суши, с которой происходит снос материала, наличием высотной поясности, влияющей на состав палиноспектров (далее в тексте ПС). Для правильного толкования результатов спорово-пыльцевого анализа важно понимать закономерности формирования ПС в донных осадках морей и учитывать степень их адекватности составу растительности побережья. Основная задача данной работы – анализ особенностей субрецентных ПС, извлеченных из поверхностного ненарушенного слоя морских осадков различных участков внешнего шельфа и континентального склона моря Лаптевых, а также в палеодолинах рек Лены, Анабара, Оленёка и Хатанги, отобранные в экспедициях АРК27-3 на НИС «Поларштерн» и Трансдрифт XXI на НИС «Виктор Буйницкий» в 2012-2013 гг. [1] и оценка доли в составе субрецентных морских ПС пыльцы и спор из разрушающихся эрозионными и абразионными процессами льдистых многолетнемерзлых толщ, широко распространенных на северо-востоке Сибири, Полученные данные обеспечивают актуалистическую основу региональных палеореконструкций.
РАЙОН ИССЛЕДОВАНИЯ
Море Лаптевых, окраинное море Северного Ледовитого океана, расположенное между полуостровом Таймыр и архипелагом Северная Земля на западе и Ново-Сибирскими островами на востоке, оказывает большое влияние на климат и окружающую среду Арктики благодаря обширному теплообмену с атмосферой летом и формированию значительной части морского арктического льда зимой. Его площадь составляет около 678 тыс. кв. км, объём воды достигает 363 тыс. куб. км, средняя глубина - 578 м. Солёность воды низкая, ледовый покров держится более 10 месяцев в году, и только частично отступает в конце лета-начале осени. Более половины морского дна относится к материковой отмели с глубиной не более 60 метров, в южной и юго-восточной части глубина не превышает 25-30 м, а в северной - превышает 1 км. Максимальная глубина 3385 м в котловине Нансена. Вдоль материкового склона на глубинах 250-300 м движутся относительно тёплые воды (до 1,5°C), формирующиеся в северной части Атлантики и достигающие моря Лаптевых за 2,5-3 года. Поверхностные течения моря образуют циклонический круговорот, который усиливается стоком р. Лены и отвлекается в сторону Северного Ледовитого океана [2]. Взаимодействие между водными массами шельфа и континентального склона, определяемое силой/направлением морских ветров и интенсивностью речного стока, вызывает сильные реверсивные течения, которые могут распространяться на внешний шельф [3].
В море Лаптевых впадают крупные реки Лена, Яна, Хатанга, Оленёк, Анабар. Бо́льшая часть речного стока (до 70%, или около 540 км3 в год) приходится на долю р. Лены [4]. Среднегодовой расход воды в дельте р. Лена колеблется от 15 500 до 17 175 м3/сек. Современная седиментационная обстановка на внутреннем шельфе определяется речным стоком, с которым поставляется огромная масса твердого и органического вещества в виде взвеси, еще и морской и речной абразией многолетнемерзлых толщ [5]. Основная масса взвеси, содержащей органику, осаждается, согласно А.П. Лисицыну [6], в маргинальном фильтре, т.е. в пределах материковой отмели (рис. 1).

Рис. 1. Космический снимок дельты р. Лена с отчетливым «следом» речной взвеси, вынесенной на шельф моря Лаптевых [4]
Климат моря Лаптевых и его побережья арктический, крайне суровый. Максимальные значения радиационного баланса приходятся на июнь-август. Среднегодовая температура воздуха −13,2--14,3°С [7]. Продолжительность безморозного периода 40-60 дней. Среднеиюльские температуры воздуха варьируют от +4° на севере дельты Лены до +8,9°С на юге. Над морем характерны сильные и устойчивые ветры (средняя скорость 5-7 м/сек), с апреля по сентябрь преимущественно северного и северо-восточного направлений. Осадков выпадает мало (172-241 мм в год), 75% приходится на апрель-октябрь. Устойчивый снежный покров на побережье образуется в конце сентября. Его средняя продолжительность - 250 дней. Разрушение начинается в последней декаде мая, полный сход - в среднем в первой декаде июня. Снегопады бывают в любое время года. Мощность слоя сезонного протаивания составляет 0,1-0,3 м под моховой дерниной и до 1 м на оголённых участках. Широко распространены мерзлотные тундровые перегнойно-глеевые и перегнойно-глееватые почвы.
Берега моря Лаптевых и побережье полуострова Таймыр занимает травяно-кустарничковая мохово-лишайниковая арктическая тундра [5]. По долинам рек и протоков встречаются лесотундровые виды, а по Хараулахским горам (северная часть Верхоянского хребта) – горные виды. К северу от о. Тит-Ары в распадках долин небольших ручьев, прорезающих северные склоны кряжа Чекановского, прилегающие к дельте р. Лены, а также на о. Самойловском встречается лиственница Larix cajanderi Mayr [8]. В комплексе с тундрами на отрицательных формах рельефа распространены тундровые болота, чаще полигонально-валиковые тундроболотные комплексы с травяными сфагновыми тундрами на валиках и травяными болотами в полигонах. Более сухие участки занимают пятнистые тундры. Кустарниковый покров слабо развит, образован Salix glauca L., S. pulchra Cham., Betula nana L. В травяно-кустарничковом ярусе (20-70%) преобладают Ledum palustre L., Vaccinium vitis-idaea L., Vaccinium uliginosum L., Cassiope tetragona (L.) D. Don, из разнотравья Rubus chamaemorus L., из злаков Arctagrostis arundinacea (Trin.) Beal., Poa sp. Встречаются Carex vesicata Meinsh., Eriophorum vaginatum L., Poa pratensis L. Рассеянно произрастают Saxifraga punctata L., S. hieracifolia Waldst. et Kit, Dryas octopetala L., Papaver lapponicum ssp..Pedicularis lapponica L., P. sceptrum-carolinum L., Stellaria angarae M. Pop., Aconogonon tripterocarpum (A. Gray) Hara., Persicaria amphibia (L.) S.F. Gray, Ranunculus lapponicus L., Arctous alpina (L.) Niedenzu, Equisetum pratense Ehrh., Petasites frigidus (L.) Fries, Saussurea alpina (L.) DC., Carex sp. Моховой покров сплошной,(80-100%) из Aulacomnium turgidum (Wahlenb.) Schwägr., Hylocomium splendens (Hedw.) Bruch et al., Tomenthypnum nitens (Hedw.) Loeske, Sphagnum sp., Aulacomnium palustre (Hedw.) Schwägr. Лишайники играют значительную роль в покрытии (5-10%), встречаются Peltigera aphthosa (L.) Willd., Dactylina arctica (Hook.) Nyl., Flavocetraria cucullata (Bellardi) Kärnefelt et Thell,Сetraria laevigata Rassads., Stereocaulon paschale (L.) Hoffm. [9]. Растительность речных долин на побережье представлена лугами и кустарниковыми сообществами. Луга в тундровой зоне связаны с экотопами, почвы которых периодически обогащаются свежими наилками, достаточно увлажнены и дренированы. Их основные площади приходятся на поймы рек, в остальных случаях они встречаются крохотными фрагментами. Наиболее часты в пойме реки Лена хвощовые и пушицево-хвощовые луга. Equisetum pratense Ehrh., E. arvense L., Eriophorum vaginatum L., E. scheuchzeri Hoppe, Arctophila fulva (Trin.) Anders., Poa pratensis L., Salix glauca L., единично отмечаются Tanacetum vulgare subsp. boreale (Fisch. ex DC.), некоторые виды осок. Кустарниковая растительность представлена зарослями ивы высотой до 0,5 м. Для арктических тундр на побережье моря Лаптевых характерны Carex aquatilis Wahlenb. ssp. stans, Eriophorum angustifolium Honck., Salix polaris Wahlenb, Salix nummularia Anderss., Cassiope tetragona (Pall.) D. Don, зеленые мхи и лишайники [7]. Для равнинных субарктических тундр типичными представителями являются Salix pulchra Cham., S. glauca L., S. reptans Rupr., Betula exilis Sukaczev, Vaccinium vitis-idaea L., Cassiope tetragona (Pall.) Dryas punctata Juz., Salix polaris Wahlenb. Среди трав господствуют Carex aquatilis Wahlenb. ssp. stans (Drejer) Hulten и Eriophorum angustifolium Honck. Моховый ярус представлен, в основном, Aulacomnium turgidum (Wahlenb.) Schwagr. и Hylocomium splendens (Hedw.) B.S.G. var. аlaskanum [7]. Рассматривая состав прибрежной растительности как источник поступления пыльцы и спор в донные отложения, отметим, что прибрежная растительность представлена практически исключительно тундровыми биомами и характеризуется низкой пыльцевой продуктивностью. Граница ареалов распространения древесной растительности Pinus s/g Diploxylon (P. sylvestris L.) и Betula sect. Albae (B. pendula Roth. + B. pubescens Ehrh.) проходят в 600-700 км от дельты р. Лены, граница распространения Picea obovata Ledeb. находится в 200 км к югу от дельты р. Лены [10].
МАТЕРИАЛ И МЕТОДЫ
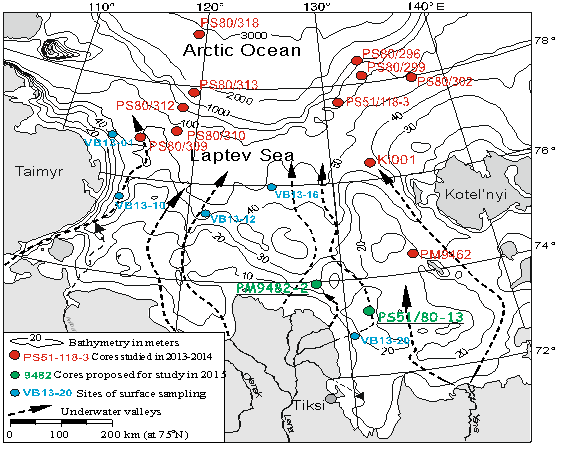
Материалом для исследования послужили образцы ненарушенного поверхностного слоя донных осадков, впервые поднятые 8 мультикорерами на внешнем шельфе и континентальном склоне моря Лаптевых в экспедиции ARK27-3 на НИС «Поларштерн» в 2012 г. и 5 мультикорерами в затопленных палеодолинах рек Лены, Анабара, Оленека и Хатанги в экспедиции Трансдрифт XXI на НИС «Виктор Буйницкий» в 2013 г. (рис. 2).
Рис. 2. Местоположение станций отбора проб поверхностных донных осадков
Основной метод исследования - маринопалинологический, который включает в себя анализ видового состава и изменений концентрации как терригенных, так и водных палиноморф. Метод анализа водных палиноморф, представленных двумя экологическими группами (морской и пресноводной), в последние годы приобретает все большее значение в практике палеоокеанологических реконструкций, поскольку позволяет реконструировать гидрологические условия морского бассейна, вариации речного стока, биопродуктивность вод и другие важнейшие экологические параметры морской среды [11, 12, 13, 14].
Для извлечения микрофоссилий из осадка использована методика В.П. Гричука [15]. Перед началом химической обработки в каждый образец осадка, высушенного предварительно методом вакуумной сушки, была добавлена таблетка с фиксированным числом окрашенных спор Lycopodium clavatum (партия № 1031) для расчета концентрации идентифицированных палиноморф [16], представленных в статье как количество микрофоссилий на 1 грамм сухого осадка. Таксономическая идентификация проведена с помощью опубликованных ключей и атласов [11, 12, 15, 17, 18, 19, 20, 21] под микроскопом Motic-B1-220A при увеличении ×400. Для идентификации микрофоссилий древнего облика и выяснения их стратиграфического распространения использовались работы с описанием ПС из отложений фанерозоя районов, относившихся к единой палеофлористической Сибирской области[22, 23]. В каждом образце с внешнего и внутреннего шельфа подсчитано не менее 170-200 зерен пыльцы; непыльцевые палиноморфы (далее в тексте НПП) и переотложенные дочетвертичные микрофоссилии подсчитаны в дополнение. В образцах с континентального склона (313, 318, 296 и 299) с трудом удалось насчитать в сумме 200 микрофоссилий, включая древние и переотложенные. В группу НПП объединены споры, органические остатки водных микроорганизмов морского (цисты динофлагеллат, акритархи, органическая часть скелета фораминифер) и пресноводного (хлоро- и харофитовые водоросли) генезиса, споры грибов рода Glomus и печеночников. Процентное соотношение отдельных пыльцевых таксонов высчитывалось от общей суммы пыльцы древесных и травянистых растений. Процентное содержание спор – от суммы пыльцы, доля переотложенных дочетвертичных спор и пыльцы – от общей суммы зарегистрированных микрофоссилий в образце. Процентные соотношения в группе НПП рассчитаны от общей суммы НПП. Для подсчета процентных соотношений и построения спорово-пыльцевых диаграмм использован пакет программ Tilia/TiliaGraph/TGView [24, 25].
РЕЗУЛЬТАТЫ
Все изученные ПС содержали микрофоссилии широкого возрастного диапазона (от современных до девонских) и различной степени сохранности. В целом, не несущая признаков переотложения часть ПС характеризуется невысоким таксономическим разнообразием, но, тем не менее, отражает состав растительности четырех ботанико-географических зон – арктической пустыни, тундры и лесотундры и тайги. Локально-региональный компонент ПС составляет пыльца приморских луговых галофитов и пойменных тундровых сообществ материкового обрамления моря Лаптевых, а также травянистых биоценозов зоны полярных пустынь Новосибирских островов и северного побережья полуострова Таймыр. В споровой части ПС этот компонент формируется, в основном, арктическими плаунами, сфагновыми и зелеными мхами. К дальнезаносной группе отнесена пыльца древесных растений лесотундровой и таежной зон и легко плавучие споры папоротников, а также отмеченная только в осадках восточной части материкового склона единичная пыльца дуба, вяза и липы.
В общем составе ПС выявлены существенные различия в зависимости от района захоронения компонентов ПС в донные осадки (рис. 3).
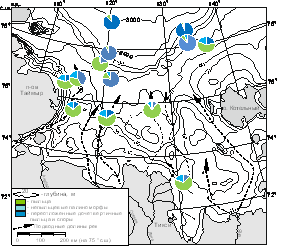
Рис. 3. Общий состав изученных ПС поверхностного слоя осадков моря Лаптевых, %
Так, основу субрецентных ПС внешнего шельфа и континентального склона составляют переотложенные пыльца и споры дочетвертичного возраста (до 94% от общего количества микрофоссилий), что свидетельствует об активном влиянии на формирование ПС береговой, склоновой и донной абразии. Их концентрация в поверхностном слое осадков западной части внешнего шельфа достигает максимальных значений более 60х103 зерен/г, в то время как в восточной – намного ниже и не превышает 23х103 зерен/г. Таксономический состав переотложения весьма разнообразен (до 35-40 таксонов в большинстве образцов). Доминирует пыльца мезозойских хвойных (Picea, Pinus, Gingko sp., Taxodiaceae/Cupressaceae), встречена пыльцаAsaccitessp., Caytoniasp., EntylissacycadiformisNaum., Cycadaceae, часто - позднемеловая-палеогеновая пыльца формальных родов (Orbiculapollisglobosus(Chlon.) Chlon., Extratriporopollenites, Nudopollis, Trudopollis), реже - WodehouseiaspinataStanley. К позднекайнозойской группе отнесена пыльца Myrica,Nyssa, Pterocarya, Carya, Ilex, Rhamnus, и пыльца берез темно-коричневого цвета с утолщенной экзиной. В группе спор наиболее часто встречались Pliciferadelicata (Bolch.) Bolch., Gleichenialaeta Bolch., G. umbonata Bolch.G. angulata Bolch., Osmunda sp., реже – Tripartina paradoxa, Dipteridaceae, Phlebopteris polypodioides, Leiotriletes. В «молодой» части ПС западной части внешнего шельфа и континентального склона встречены лишь единичная пыльца сосны сибирской, злаков и плавучих спор папоротников (рис. 4, А).

Рис.4. Генерализованный таксономический состав субфоссильных ПС в поверхностном слое донных осадков, %: А – внешнего шельфа и континентального склона; Б – внутреннего шельфа. АР (arboreal pollen) – пыльца древесных и кустарниковых растений; NAP (non-arboreal pollen) – пыльца трав и кустарничков
Несмотря на удаленность точек пробоотбора на сотни километров от берега, в ПС из поверхностных осадков района к востоку от 135º в.д., доминирует пыльца растений, широко распространенных в пойменных биоценозах дельты Лены [26, 27, 28] - Alnаsterfruticosus, Cyperaceae, Poaceae и Betulanana-type, в ассоциации со спорами тундровых растений и значительным участием колониальных пресноводных водорослей родов Pediastrum и Botryococcus(в сумме до 14% от общего числа зарегистрированных микрофоссилий).
В ПС из поверхностных осадков внутреннего шельфа преобладает пыльца современных растений материкового обрамления моря Лаптевых, ее сохранность весьма удовлетворительна, цвет – зеленоватый, а в некоторых зернах (преимущественно, берез) сохранено внутренне содержимое. Доля переотложенных дочетвертичных микрофоссилий в ПС внутреннего шельфа не превышает 9-16% (рис. 4, Б). Основу ПС формируют всего несколько таксонов: дальнезаносная пыльца сосны подродов Haploxylon и Diploxylon, а также локально-региональные компоненты - пыльца кустарничковых берез, злаков и кустарниковой ольхи. Определена также пыльца растений, характерных для луговых сообществ Сибирской Арктики (Rosaceae, Ranunculaceae и Polygonaceae).
Концентрация микрофоссилий варьирует в пределах от 18.1×103 зерен/г в центральной части внешнего шельфа до 135×103 зерен/г в восточной части внешнего и юго-восточной части внутреннего шельфа, примыкающей к дельте Лены. В направлении континентального склона она уменьшается до 2-5×103 зерен/г.
В группе микрофитопланктона внутреннего шельфа, особенно в поверхностном слое осадков палеодолин рек Хатанги и Лены (ст. 302 и 309, см. рис. 2), доминируют пресноводные колониальные водоросли рода Pediastrum. концентрация которых достигает 7,6-16,5×103 зерен/г, а суммарная доля в общей численности НПП – от 11 до 57%. В обедненной ассоциации диноцист в осадках внутреннего шельфа доминируют холодноводные гетеротрофные виды Islandiniumvar.minutum и Brigantedinium spp., способные выдерживать низкую соленость прибрежных вод [11, 12, 13]. В ПС континентального склона единично отмечены автотрофные виды Spiniferiteselongatus, Nematosphaeropsislabyrinthus и Operculodiniumcentrocarpum. В восточной части континентального склона в значительных количествах (до 40% от общей суммы НПП) и в концентрации, превышающей 5×103 зерен/г, зарегистрирован Pediastrumkawraiskii.
ОБСУЖДЕНИЕ И ВЫВОДЫ
Выявленные различия в составе ПС из поверхностных осадков различных участков шельфа и континентального склона моря Лаптевых и их значительное сходство с пыльцевыми записями, полученными Л.А. Савельевой и Е.А. Рашке, изучавшими состав пыльцы и спор в осадках, слагающих различные геоморфологические уровни дельты реки Лены [26, 27]. позволяют сделать вывод о господствующей роли речного стока в транспортировке пыльцы и спор на шельф моря Лаптевых и о подчиненной роли фактора воздушного переноса, доминирующего, например, в Баренцевом море [28]. И действительно, в летне-весенней розе ветров над морем Лаптевых преобладают ветры северо-восточного направления [29], т.е. из высокоширотных районов Арктики, где на островах обитают растения с низкой пыльцевой продуктивностью. По этой причине доля пыльцы и спор, занесенных ветром из южных районов, в том числе и из таежной зоны, в ПС донных осадков, видимо, невысока. Значительно бóльшая концентрация пыльцы и пресноводных зеленых водорослей в осадках восточной части континентального склона, скорее всего, объясняется последующим перераспределением органической взвеси по площади бассейна придонной циркуляцией водных масс. Изменения концентрации пыльцы, спор и пресноводных зеленых водорослей могут свидетельствовать об изменениях интенсивность и объема речного стока.
Для оценки возможных источников поступления пыльцы и спор на шельф моря Лаптевых нам также представляется необходимым рассмотреть состав ПС в едомных отложениях и повторно-жильных льдах, широко распространенных в Сибирской Арктике [30, 31, 32, 33]. На побережье моря Лаптевых и в районах, дренируемых реками бассейна моря Лаптевых высокольдистые синкриогенные отложения с повторно-жильными льдами занимают огромные площади. В обнажениях ледового комплекса на побережье моря Лаптевых и в дельте р.Лена как правило выделяются три основных льдонасыщенных макроцикла. Синкриогенные едомные отложения, вмещающие ледяные жилы, характеризуются линзовидной сетчатой криогенной текстурой. Повторно-жильные могут превышать 3 м по ширине и до 40 по высоте Содержание льда в межполигональных блоках составляет от 80 до 180% (отношение веса льда к весу сухой породы). Горизонты палеопочв обогащены органическим веществом и в них содержится меньше льда по сравнению с минеральными прослоями и с горизонтами. Общее объемное содержание льда в ледовом комплексе может превышать 80%. Органика в едомных отложениях хорошо сохраняется она представлена автохтоным торфом, макроостатками насекомых и растений, костями млекопитающих - мамонтов, лошадей, бизонов и грызунов. Едомные отложения содержат пыльцу и споры различной степени сохранности, которые при разрушении этих отложений речной эрозией и абразией, логично предположить, выносятся речными водами в море в составе твердой взвеси, наряду с диатомовыми и зелеными водорослями. Пыльца и споры в едомных отложениях Сибирской Арктики представлены 38 палинотаксонами и 5 таксонами непыльцевых палиноморф [30, 33, 34 35, 36, 37]. Содержание пыльцы трав и кустарничков в большинстве образцов едомных отложений в возрастном интервале 40-10 тыс. лет назад характеризуется низкой концентрацией пыльцы и доминированием пыльцы травянистых таксонов (до 80%). Обычные доминанты в едомных отложениях – пыльца Cyperaceae (20-80%) и Роасеае (20-60%). Суммарная доля пыльцы доминирующих тундровых кустарников Betula nana-type, Salix sp. и Alnaster fruticosus обычно не превышает 20% от общего количества пыльцы, при этом содержание пыльцы Salix в отдельных горизонтах достигает 4%. Пыльца Artemisia sp., как правило, встречается в количестве до 15%. Среди трав доминирует пыльца Brassicaceae (до 8%), Caryophillaceae (до 15%), Ranunculaceae (до 15%) и Papaveraceae (до 5%). Споры Sphagnum, Selaginella rupestris, Lycopodium, Huperzia встречаются часто, но в низкой концентрации. Cодержание спор колеблется от 1-2 до 40%, обычно наиболее высока доля спор Selaginella rupestris [31]. Палиноспектры повторно-жильных льдов едомы Быковского полуострова представлены 34 палинотаксонами, характеризуются преобладанием пыльцы кустарников Betula nana-type и Alnaster fruticosus (12-56%), встречается пыльца Pinus s/g Haploxylon (до 20%) , среди трави кустарничков абсолютно доминирует пыльца разнтравья (36-40%) среди пыльцы трав доминирует пыльца Caryophillaceae (до 15%), Ranunculaceae (до 12%) и Papaveraceae (до 3%), с заметным участием пыльцы Artemisia (8-28%), в споровой части спектра отмечены споры Sphagnum sp. (10-12%) а также плаунов Lycopodium, Huperzia (8-12) [34]. В едомах Мамонтов Клык, Ойгосский Яр, Буор-Хая, Чекановский мост, Курунгнах, Быковский полуостров описаны НПП - водоросли Zignemataceae, Botryоcoccus, споры грибов Glomus, Microthyrium, остатки тихоходок Macrobiothus [35, 36]. Данные таксоны, вполне вероятно, могут быть показателем участия палиноморф из едомных отложений в ПС донных осадков шельфа моря Лаптевых. Мы допускаем, что часть пыльцы тундровых кустарников, а также спор Sphagnum и Selaginella rupestris, зарегистрированныхв ПС поверхностных осадков моря Лаптевых, могла быть вымыта из едомных отложений, а, следовательно, должна быть отнесена к переотложенной части ПС, но отделить данную часть от остальной пыльцы не представляется возможным. В основании разрезов едомы на побережье моря Лаптевых и в дельте р. Лена прослеживается горизонт с высоким содержанием (до 40%) пыльцы Pinaceae очень темного цвета, значительными повреждениями экзины и различной степенью автофлюоресценции. Горизонт датируется возрастом древнее 49 тыс. лет [33, 34]. Вероятно, определенная доля пыльцы Pinaceae с похожими внешними признаками в ПС поверхностных осадков шельфа и континентального склона могла быть вынесена на шельф из подстилающего едомные отложения горизонта в процессе речной эрозии и береговой абразии.
Палиноиндикационные исследования поверхностных осадков моря Лаптевых позволили выявить важную закономерность в распределении по площади изучаемого региона единичной пыльцы «экзотических», нехарактерных для Арктики широколиственных растений. Исследованиями прошлых лет установлено, что некоторые морфологические свойства пыльцы широколиственных пород деревьев (вяза, дуба, липы) препятствуют ее вовлечению в дальний воздушный перенос, поэтому она фоссилизуется в пределах ареала продуцирующих растений [30, 37, 38]. При этом, водный занос реками рассматривается как наиболее вероятный источник ее транспортировки на большие расстояния от ареала. В изучаемом регионе без признаков переотложения она идентифицирована лишь в пробах осадков восточной части континентального склона, на расстоянии более 600 км от берега и запредельно далеко от границ ареала продуцирующих ее растений, расположенного вне Восточной Сибири. Вероятно, данная пыльца вовлекается в водный перенос в западноевропейском регионе и поступает в море Лаптевых с трансформированными водами атлантического происхождения (АТВ), движущимися вдоль континентального склона, таким образом, маркируя зону распространения АТВ. Наши выводы согласуются с данными, полученными по Баренцевоморскому региону [28]. В поверхностных осадках шельфа Баренцева моря пыльца широколиственных деревьев в количестве до 2-6% присутствует повсеместно на Мурманском и Северо-Канинском мелководьях, в меньшем количестве – в осадках Южно-Новоземельского желоба, и отсутствует в осадках закрытой со всех сторон сушей Чёшской губе, Центральной впадине, Горле Белого моря и шельфа Шпицбергена. В группе водных палиноморф индикатором влияния АТВ на арктические экосистемы принято считать цисты автотрофных видов динофлагеллат [11, 12, 13], в том числе Spiniferiteselongatus, Nematosphaeropsislabyrinthus и Operculodiniumcentrocarpum, намивстреченные только в осадках западной и центральной частей внешнего шельфа и континентального склона.
Присутствие пыльцы приморских луговых галофитов, предпочитающих засоленные почвы лайд (Artemisia, Asteraceae, Poaceae и Chenopodiaceae), в количествах, более единичного, может использоваться для оценки близости береговой линии. Субрецентные ПС внутреннего шельфа гораздо ближе по составу к субрецентным ПС, формирующимися на заливаемых половодными водами элементах рельефа, т.е. к палиноспектрам пойм.
Анализ изменений процентного соотношения между основными группами водных палиноморф по изученной площади показывает повсеместное господство пресноводных зеленых водорослей над морскими цистами динофлагеллат как в палеодолинах рек, так и в восточной части континентального склона в зоне влияния речного стока Лены.
Исследование выполнено в рамках стипендиальной программы российско-германской Лаборатории по морским и полярным исследованиям им. О.Ю. Шмидта (ААНИИ, г. Санкт-Петербург), и грантов: РФФИ №15-05-08497 и РФФИ № 17-05-00794.
Библиография
1. Руденко О.В., Енина В.В., Овсепян Я.С. Вариации состава субфоссильных палиноспектров в донных осадках шельфа и континентального склона моря Лаптевых //Мат-лы XХI Междунар. научн. конф. (Школы) по морской геологии. Москва, 16-20 ноября 2015 г. М.: ГЕОС, 2015. Т.1. С. 218-221.
2. Gordeev V.V., Martin J.M., Sidorov I.S., Sidorova M.V. A reassessment of the Eurasian river input of water, sediment, major elements, and nutrients to the Arctic Ocean // American Journal of Sciences. 1996. V.296. Pp. 664-691.
3. Dmitrenko I.A., Hoelemann J.A., Kirilov S.A., Wegner C., Gribanov V.A., Berezovskaya S.L., Kassens H. Thermal regime of the Laptev Sea bottom layer and affecting processes // Earth’s Cryosphere. 2001 V. 5 (3). 2001. Pp. 40-55.
4. http://www.fesk.ru/wetlands
5. Bauch H.A., Kassens H., Kunz-Pirrung M., Naidina O., Thiede J. Composition and flux of Holocene sediments on the eastern Laptev Sea shelf. Arctic Siberia // Quaternary Research 55. 2001. Р. 344-351.
6. Lisitzin, A.P. Marginal filter in the oceans // Oceanology. 1995. V. 34 (5). P. 671–682.
7. Атлас Арктики. 1985. М.: ГУГК.
8. Исаев А.П., Габышева Л.П., Михалева Л.Г., Соломонов Н.Г. Растительные сообщества острова Тит-Ары (Восточная Сибирь) // Современные проблемы науки и образования. 2016. № 6. URL: https://science-education.ru/ru/article/view?id=25695 (дата обращения: 17.10.2017).
9. Егорова А.А. Растительный покров Усть-Ленского заповедника // Заповедники Российской Арктики: проблемы и пути их решения. Материалы Всероссийской научно-практической конференции, посв. 25-летию Госуд. прир. запов. «Усть-Ленский» (п. Тикси Респ. Саха (Якутия), декабрь 2010 г.). 2010. С. 49-50.
10. Соколов С. Я., Связева О. А., Кубли В. А., Скворцов А. К., Грудзинская И.А., Огуреева Г.Н. Ареалы деревьев и кустарников СССР. В трех томах. Т. 1. Л.: Наука. 1977. 164 с.
11. Matthiessen J.,. Distribution patterns of dinoflagellate cysts and other organic-walled microfossils in recent Norwegian-Greenland Sea sediments // Marine Micropaleontology. 1995 V. 24. P. 307 – 334.
12. Kunz-Pirrung M. Aquatic palynomorphs: reconstruction of Holocene seasurface water masses in the eastern Laptev Sea // Berichte Polarforschung. 1998. V. 281, 117p.
13. Polyakova, Ye.I., Bauch, H.A., Klyuvitkina, T.S. Early to middle Holocene changes in Laptev Sea water masses deduced from diatom and aquatic palynomorph assemblages // Global and Planetary Change. 2005. V. 48. P. 208-222.
14. Rudenko O., Tarasov P.E., Bauch H.A., Taldenkova E.E., A Holocene palynological record from the northeastern Laptev Sea and its implications for palaeoenvironmental research // Quaternary International. 2014. V. 348. P. 82-92.
15. Пыльцевой анализ / Под ред. И.М. Покровской. М.: Госгеолиздат. 1950.
16. Stockmarr J. Tablet spores used in absolute pollen analysis // Pollen Spores. 1971. V. 13. P.616-621.
17. Komárek J., Jankovská V. Review of the green algal genus Pediastrum: Implication for pollen analytical research. Bibliotheca Phycologica. 2001. V. 108. P. 1-127.
18. Куприянова Л.А. Палинология сережкоцветных. М.: Наука. 1965. 213 с.
19. Куприянова Л.А., Алешина Л.А. Пыльца и споры растений флоры СССР. Т. 1. Л.: Наука, 1972. 171 с.
20. Савельева Л.А., Рашке Е.А., Титова Д.В. Атлас фотографий растений и пыльцы дельты реки Лены. СПб. 2013. 114 с.
21. Соколовская А.П. пыльца растений Арктики. Л.: Научный бюллетень ЛГУ. Т. 33. 1955. С. 244-292.
22. Атлас нижнемеловых спорово-пыльцевых комплексов некоторых районов СССР / Под ред. И.М. Покровской, Н.К. Стельмак. М.: Недра, 1964. 551 с.
23. Палеопалинология / Под ред. И.М. Покровской. Л.: Недра, 1966. Т.1-3.
24. Grimm, E.C. TILIAGRAPH vl.25 (computer software). Illinois State Museum, Research and Collections Center, Springfield, IL, USA. Google Scholar. 1991.
25. Grimm, E.C. TGView 2.0.2 (Software). Illinois State Museum, Research and Collections Center, Springfield, IL. 2004.
26. Савельева Л.А., Морозова Е.А., Большиянов Д.Ю., Титова Д.В., Вахрамеева П.С. Первые результаты исследования спорово-пыльцевых спектров поверхностных проб в дельте реки Лены // Заповедники Российской Арктика: проблемы и пути их решения. Материалы Всероссийской научно-практической конференции, посвященной 25-летию Государственного природного заповедника «Усть-Ленский» (п. Тикси, Республика Саха (Якутия)). 2010. С. 117-121.
27. Рашке Е.А., Савельева Л.А. Субрецентные спорово-пыльцевые спектры и современная растительность дельты реки Лены, Российская Арктика // Сибирский экологический журнал. Новосибирск: Издательство СО РАН. 2017. №4.
28. Руденко О.В., Полякова Е.И. Особенности формирования спорово-пыльцевых спектров в поверхностных осадках Баренцева моря //Седиментологические процессы и эволюция морских экосистем в условиях морского перигляциала. Апатиты: Изд-во КНЦ РАН. 2001. С.111-120.
29. Gervais M., Atallah E.H., Gyakum J.R., Tremblay B. Arctic Air Masses in a Warming World // Journal of Climate 29(7). 2016. DOI: 10.1175/JCLI-D-15-0499.1
30. Васильчук А.К. Особенности формирования палиноспектров в криолитозоне России. М.: Изд-во Моск ун-та. 2005. 245 с.
31. Украинцева В.В., Поспелов И.Н. О связях состава растительности и состава спорово-пыльцевых спектров поверхностных проб (устье р. Оленья, полуостров Таймыр) // Известия РАН. Серия Географическая. 2006. № 3. C. 97-109.
32. Васильчук А.К. Региональная и дальнезаносная пыльца в тундровых палиноспектрах // Известия Российской Академии Наук. Серия биологическая. № 1. 2005. С. 85–99. (Vasil'chuk A. Regional and extra-local pollen in tundra pollen samples // Biology Bulletin of the Russian Academy of Sciences. 2005. V. 32. N 1. P. 75–84.)
33. Васильчук А.К. Палинология и хронология полигонально-жильных комплексов в криолитозоне России (Palynology and chronology of polygonal ice wedge complexess in Russia permafrost area) / Под редакцией действительного члена РАЕН, профессора Ю.К.Васильчука. М.: Изд-во Моск. ун-та. 2007. 488 с.
34. Васильчук Ю.К. Изотопно-кислородный состав подземных льдов (опыт палеогеокриологических реконструкций). Oxygen Isotope Composition of Ground Ice (Application to Paleogeocryological Reconstructions). М.: Изд. Отдел. Теоретических проблем РАН. МГУ, ПНИИИС. 1992. В 2–х томах. Т.1. – 420 с. Т.2 – 264 с.
35. Schirrmeister L., Schwamborn G., Overduin P.P., Strauss J, Fuchs M.C., Grigoriev M., Yakshina I., Rethemeyer J., Dietze E., Wetterich S. Yedoma Ice Complex of the Buor Khaya Peninsula (southern Laptev Sea) // Biogeosciences. 2017. V. 14. Iss. 5. P. 1261-1283.
36. Wetterich S., Tumskoy V., Rudaya N., Andreev A.A., Opel T., Meyer H., Schirrmeister L., Hüls M. Ice Complex formation in arctic East Siberia during the MIS3 Interstadial // Quaternary Science Reviews. 2014. V. 84. P. 39-55.
37. Wetterich S., Kuzmina S., Andreev A.A., Kienast F., Meyer H., Schirrmeister L., Kuznetsova T., Sierralta M. Palaeoenvironmental dynamics inferred from late Quaternary permafrost deposits on Kurungnakh island, Lena delta, Northeast Siberia, Russia // Quaternary Science Reviews. 2008. V. 27. P. 1523–1540.
References
1. Rudenko O.V., Enina V.V., Ovsepyan Ya.S. Variatsii sostava subfossil'nykh palinospektrov v donnykh osadkakh shel'fa i kontinental'nogo sklona morya Laptevykh //Mat-ly XKhI Mezhdunar. nauchn. konf. (Shkoly) po morskoi geologii. Moskva, 16-20 noyabrya 2015 g. M.: GEOS, 2015. T.1. S. 218-221.
2. Gordeev V.V., Martin J.M., Sidorov I.S., Sidorova M.V. A reassessment of the Eurasian river input of water, sediment, major elements, and nutrients to the Arctic Ocean // American Journal of Sciences. 1996. V.296. Pp. 664-691.
3. Dmitrenko I.A., Hoelemann J.A., Kirilov S.A., Wegner C., Gribanov V.A., Berezovskaya S.L., Kassens H. Thermal regime of the Laptev Sea bottom layer and affecting processes // Earth’s Cryosphere. 2001 V. 5 (3). 2001. Pp. 40-55.
4. http://www.fesk.ru/wetlands
5. Bauch H.A., Kassens H., Kunz-Pirrung M., Naidina O., Thiede J. Composition and flux of Holocene sediments on the eastern Laptev Sea shelf. Arctic Siberia // Quaternary Research 55. 2001. R. 344-351.
6. Lisitzin, A.P. Marginal filter in the oceans // Oceanology. 1995. V. 34 (5). P. 671–682.
7. Atlas Arktiki. 1985. M.: GUGK.
8. Isaev A.P., Gabysheva L.P., Mikhaleva L.G., Solomonov N.G. Rastitel'nye soobshchestva ostrova Tit-Ary (Vostochnaya Sibir') // Sovremennye problemy nauki i obrazovaniya. 2016. № 6. URL: https://science-education.ru/ru/article/view?id=25695 (data obrashcheniya: 17.10.2017).
9. Egorova A.A. Rastitel'nyi pokrov Ust'-Lenskogo zapovednika // Zapovedniki Rossiiskoi Arktiki: problemy i puti ikh resheniya. Materialy Vserossiiskoi nauchno-prakticheskoi konferentsii, posv. 25-letiyu Gosud. prir. zapov. «Ust'-Lenskii» (p. Tiksi Resp. Sakha (Yakutiya), dekabr' 2010 g.). 2010. S. 49-50.
10. Sokolov S. Ya., Svyazeva O. A., Kubli V. A., Skvortsov A. K., Grudzinskaya I.A., Ogureeva G.N. Arealy derev'ev i kustarnikov SSSR. V trekh tomakh. T. 1. L.: Nauka. 1977. 164 s.
11. Matthiessen J.,. Distribution patterns of dinoflagellate cysts and other organic-walled microfossils in recent Norwegian-Greenland Sea sediments // Marine Micropaleontology. 1995 V. 24. P. 307 – 334.
12. Kunz-Pirrung M. Aquatic palynomorphs: reconstruction of Holocene seasurface water masses in the eastern Laptev Sea // Berichte Polarforschung. 1998. V. 281, 117p.
13. Polyakova, Ye.I., Bauch, H.A., Klyuvitkina, T.S. Early to middle Holocene changes in Laptev Sea water masses deduced from diatom and aquatic palynomorph assemblages // Global and Planetary Change. 2005. V. 48. P. 208-222.
14. Rudenko O., Tarasov P.E., Bauch H.A., Taldenkova E.E., A Holocene palynological record from the northeastern Laptev Sea and its implications for palaeoenvironmental research // Quaternary International. 2014. V. 348. P. 82-92.
15. Pyl'tsevoi analiz / Pod red. I.M. Pokrovskoi. M.: Gosgeolizdat. 1950.
16. Stockmarr J. Tablet spores used in absolute pollen analysis // Pollen Spores. 1971. V. 13. P.616-621.
17. Komárek J., Jankovská V. Review of the green algal genus Pediastrum: Implication for pollen analytical research. Bibliotheca Phycologica. 2001. V. 108. P. 1-127.
18. Kupriyanova L.A. Palinologiya serezhkotsvetnykh. M.: Nauka. 1965. 213 s.
19. Kupriyanova L.A., Aleshina L.A. Pyl'tsa i spory rastenii flory SSSR. T. 1. L.: Nauka, 1972. 171 s.
20. Savel'eva L.A., Rashke E.A., Titova D.V. Atlas fotografii rastenii i pyl'tsy del'ty reki Leny. SPb. 2013. 114 s.
21. Sokolovskaya A.P. pyl'tsa rastenii Arktiki. L.: Nauchnyi byulleten' LGU. T. 33. 1955. S. 244-292.
22. Atlas nizhnemelovykh sporovo-pyl'tsevykh kompleksov nekotorykh raionov SSSR / Pod red. I.M. Pokrovskoi, N.K. Stel'mak. M.: Nedra, 1964. 551 s.
23. Paleopalinologiya / Pod red. I.M. Pokrovskoi. L.: Nedra, 1966. T.1-3.
24. Grimm, E.C. TILIAGRAPH vl.25 (computer software). Illinois State Museum, Research and Collections Center, Springfield, IL, USA. Google Scholar. 1991.
25. Grimm, E.C. TGView 2.0.2 (Software). Illinois State Museum, Research and Collections Center, Springfield, IL. 2004.
26. Savel'eva L.A., Morozova E.A., Bol'shiyanov D.Yu., Titova D.V., Vakhrameeva P.S. Pervye rezul'taty issledovaniya sporovo-pyl'tsevykh spektrov poverkhnostnykh prob v del'te reki Leny // Zapovedniki Rossiiskoi Arktika: problemy i puti ikh resheniya. Materialy Vserossiiskoi nauchno-prakticheskoi konferentsii, posvyashchennoi 25-letiyu Gosudarstvennogo prirodnogo zapovednika «Ust'-Lenskii» (p. Tiksi, Respublika Sakha (Yakutiya)). 2010. S. 117-121.
27. Rashke E.A., Savel'eva L.A. Subretsentnye sporovo-pyl'tsevye spektry i sovremennaya rastitel'nost' del'ty reki Leny, Rossiiskaya Arktika // Sibirskii ekologicheskii zhurnal. Novosibirsk: Izdatel'stvo SO RAN. 2017. №4.
28. Rudenko O.V., Polyakova E.I. Osobennosti formirovaniya sporovo-pyl'tsevykh spektrov v poverkhnostnykh osadkakh Barentseva morya //Sedimentologicheskie protsessy i evolyutsiya morskikh ekosistem v usloviyakh morskogo periglyatsiala. Apatity: Izd-vo KNTs RAN. 2001. S.111-120.
29. Gervais M., Atallah E.H., Gyakum J.R., Tremblay B. Arctic Air Masses in a Warming World // Journal of Climate 29(7). 2016. DOI: 10.1175/JCLI-D-15-0499.1
30. Vasil'chuk A.K. Osobennosti formirovaniya palinospektrov v kriolitozone Rossii. M.: Izd-vo Mosk un-ta. 2005. 245 s.
31. Ukraintseva V.V., Pospelov I.N. O svyazyakh sostava rastitel'nosti i sostava sporovo-pyl'tsevykh spektrov poverkhnostnykh prob (ust'e r. Olen'ya, poluostrov Taimyr) // Izvestiya RAN. Seriya Geograficheskaya. 2006. № 3. C. 97-109.
32. Vasil'chuk A.K. Regional'naya i dal'nezanosnaya pyl'tsa v tundrovykh palinospektrakh // Izvestiya Rossiiskoi Akademii Nauk. Seriya biologicheskaya. № 1. 2005. S. 85–99. (Vasil'chuk A. Regional and extra-local pollen in tundra pollen samples // Biology Bulletin of the Russian Academy of Sciences. 2005. V. 32. N 1. P. 75–84.)
33. Vasil'chuk A.K. Palinologiya i khronologiya poligonal'no-zhil'nykh kompleksov v kriolitozone Rossii (Palynology and chronology of polygonal ice wedge complexess in Russia permafrost area) / Pod redaktsiei deistvitel'nogo chlena RAEN, professora Yu.K.Vasil'chuka. M.: Izd-vo Mosk. un-ta. 2007. 488 s.
34. Vasil'chuk Yu.K. Izotopno-kislorodnyi sostav podzemnykh l'dov (opyt paleogeokriologicheskikh rekonstruktsii). Oxygen Isotope Composition of Ground Ice (Application to Paleogeocryological Reconstructions). M.: Izd. Otdel. Teoreticheskikh problem RAN. MGU, PNIIIS. 1992. V 2–kh tomakh. T.1. – 420 s. T.2 – 264 s.
35. Schirrmeister L., Schwamborn G., Overduin P.P., Strauss J, Fuchs M.C., Grigoriev M., Yakshina I., Rethemeyer J., Dietze E., Wetterich S. Yedoma Ice Complex of the Buor Khaya Peninsula (southern Laptev Sea) // Biogeosciences. 2017. V. 14. Iss. 5. P. 1261-1283.
36. Wetterich S., Tumskoy V., Rudaya N., Andreev A.A., Opel T., Meyer H., Schirrmeister L., Hüls M. Ice Complex formation in arctic East Siberia during the MIS3 Interstadial // Quaternary Science Reviews. 2014. V. 84. P. 39-55.
37. Wetterich S., Kuzmina S., Andreev A.A., Kienast F., Meyer H., Schirrmeister L., Kuznetsova T., Sierralta M. Palaeoenvironmental dynamics inferred from late Quaternary permafrost deposits on Kurungnakh island, Lena delta, Northeast Siberia, Russia // Quaternary Science Reviews. 2008. V. 27. P. 1523–1540.
|
 Рус
Рус